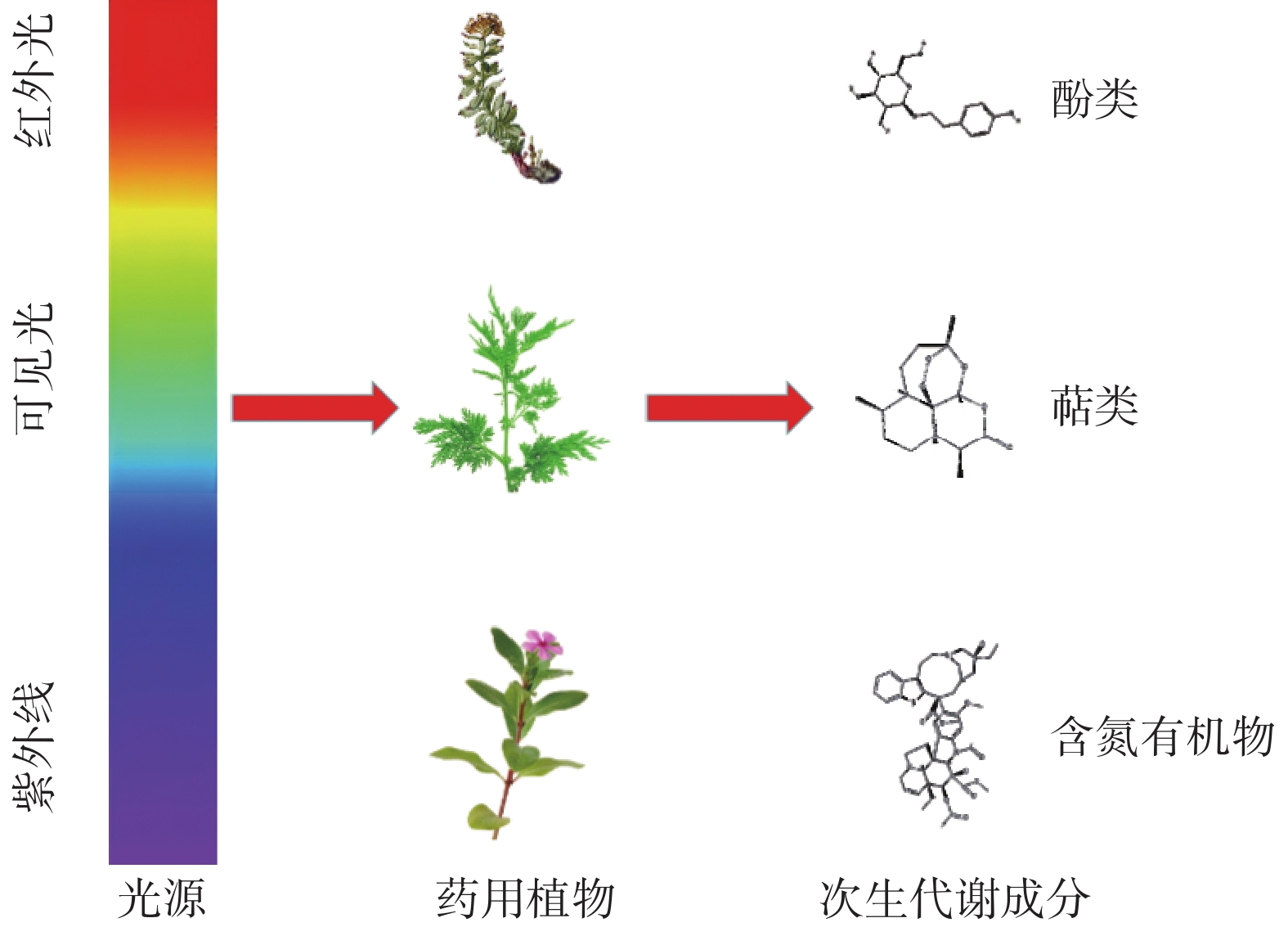


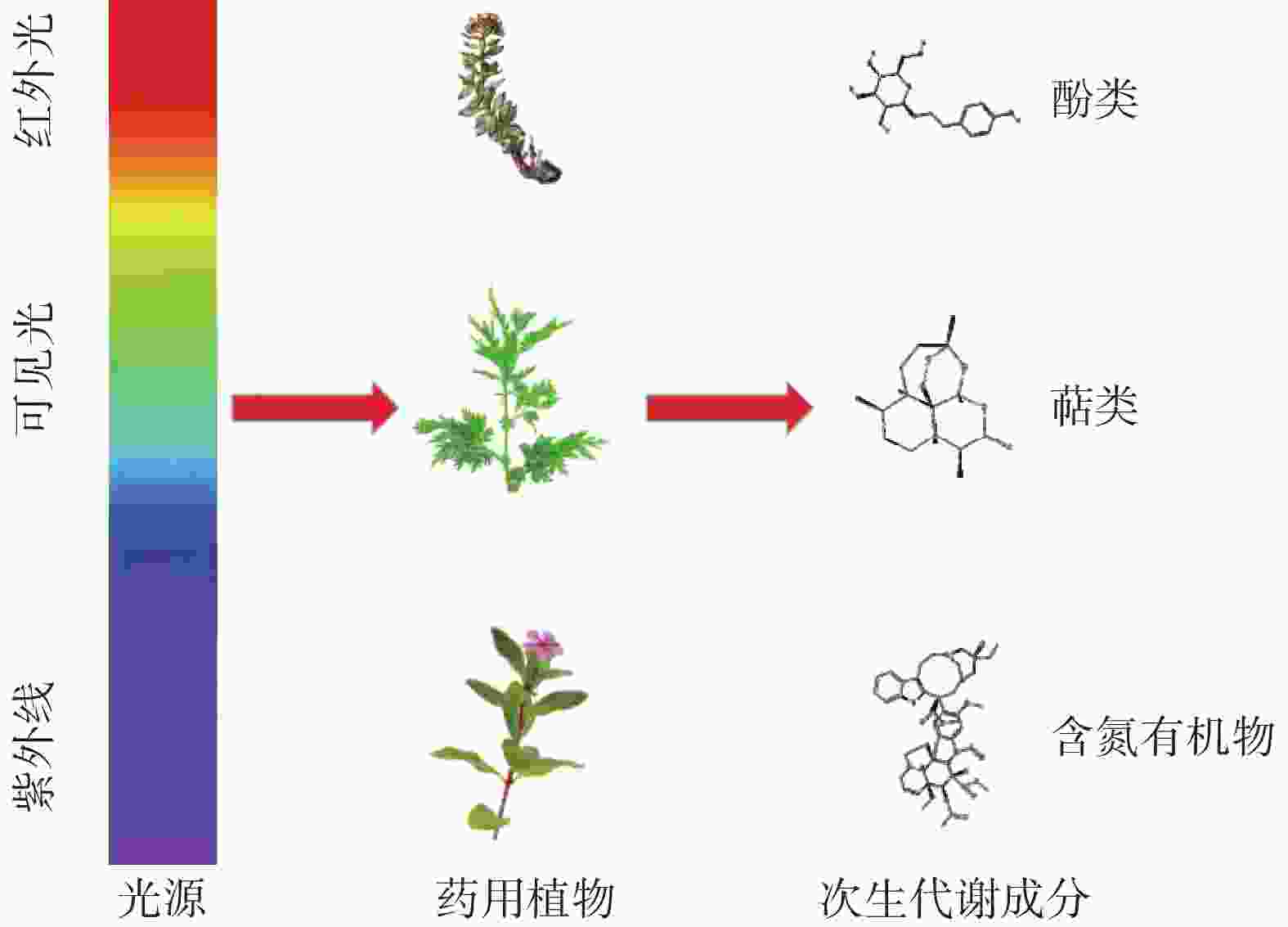
-
药用植物自古以来便是人们预防和治疗疾病的主要武器,对人类健康的发展起着举足轻重的作用。根据世界卫生组织(WHO)的数据,全球至少有25%的药物直接或间接来源于药用植物,中药有95%以上来源于药用植物,并且超过60%的抗癌药物是直接或间接从药用植物中发现的[1],所以药用植物是药物的主要来源。次生代谢成分(secondary metabolites,SMs)是指不直接参与植物生长发育,但对植物的长期生存有着至关重要作用的小分子有机化合物[2]。根据其结构和生物合成途径,植物SMs主要分为多酚类、萜类和含氮有机物,这些化合物具有抗氧化、抗炎、清除自由基、抗菌和抗病毒等活性[3]。SMs实际上是植物在长期进化中为对抗生物和非生物胁迫而促进环境适应的结果[4]。SMs的合成和积累是非常复杂的,受多种因素的影响,包括内部发育遗传回路(受调控的基因、酶)和外部环境因素(光、温度、水、盐度等)[5]。在这些因素中,光几乎影响所有类型SMs的积累(见图1),光质、光强和光周期均影响药用植物SMs的含量[6]。药用植物的SMs是临床疗效的物质基础,是评价药材质量的重要指标。光如何影响SMs生物合成对于提高药用植物SMs的产量和质量至关重要。本文就近年来国内外关于光对药用植物SMs的研究现状进行综述,以期为相关研究及药用植物高产优质栽培提供指导。
-
根据光的波长,太阳光谱可分为:紫外光(UV,100~400 nm,其中:A波段,315~400 nm;B波段,280~315 nm;C波段,100~280 nm)、可见光或光合有效辐射(PAR;蓝光:400~500 nm;绿光:500~600 nm;红光:600~700 nm)和近红外光(700~800 nm)[7]。光感受器是植物光感觉和光信号转导不可或缺的部分,植物对光的反应能力是由能够感知不同光照强度、光质、光照方向和光周期的光感受器所决定的[3]。目前在模式植物拟南芥中至少鉴定出了五类光感受器:①光敏色素(phyA–phyE),主要感知红光和红外光;②隐色素(CRY1/CRY2),主要感知蓝光和紫外A波段(UV-A);③向光素(phots),主要感知蓝光;④蓝光受体ZTLs(zeitlupes),主要感知蓝光和绿光(450~520 nm);⑤紫外线感光器也称UV抗性位点8(UVR8),主要感知紫外B波段(UV-B)[8]。光感受器通过调节特定基因的表达来刺激SMs的生化途径,SMs的生物合成与光感受器吸收的波长密切相关。
-
紫外线的波长仅占到达地球表面的太阳辐射的一小部分,但它对包括植物在内的现存物种有重大的生物影响。因此,UV可作为一种重要的非生物激发剂用于促进各种植物培养中SMs的生产[9]。
-
SMs的产生是对UV暴露增强的一种适应机制。花青素为生物类黄酮物质,而黄酮类物质最主要的功能是自由基清除能力和抗氧化能力。有研究证明:花青素是当今人类发现最有效的抗氧化剂,也是最强效的自由基清除剂,花青素的抗氧化性能比维生素C高20倍,比维生素E高50倍,并且可以吸收多余的可见光和帮助植物抵抗紫外线,清除氧自由基保护植物,是植物天然的光保护剂[10]。研究发现UV-A可促进红花檵木愈伤组织花色素和黄酮类物质的合成[11],也可以明显促进津田芜菁膨大肉质根表皮花青素的积累[12]。
-
植物通过发展保护结构和机制来应对UV-B的过度损伤。前者包括毛发、蜡和其他提供光学保护的细胞修饰,而后者涉及抗氧化酶的诱导和保护分子浓度的提高[13]。SMs的形成和产生可增强植物对UV-B的适应性,保护植物免受UV-B的损伤。
在UV-B胁迫下研究最广泛的一类生物碱是长春花体外细胞培养中的具有抗癌活性的萜类吲哚生物碱(TIAs):长春花碱和长春碱。研究表明UV-B对长春花细胞悬浮液照射5 min可将长春花碱和长春碱的产量分别提高到3倍和12倍[14]。
UV-B对萜类SMs的影响较大。紫杉醇是红豆杉属植物具有抗癌作用的二萜类化合物,研究表明,UV-B辐射(3 W/m2)12 h和24 h,可显著诱导东北红豆杉组培苗中紫杉醇的积累[15]。青蒿素是一种含有过氧化物桥结构的倍半萜内酯,在UV-B照射2 h后ADS、CPR和DBR2基因的表达被诱导,4 h和6 h后达到高峰[16]。甘草酸是甘草的主要活性成分之一,UV-B低强度(0.43 W/m2)和高强度(1.43 W/m2)照射均能提高3月龄盆栽甘草根组织中甘草酸的浓度,为对照的1.5倍,且3~6个月生盆栽植物的甘草酸浓度与3~4年生田间植物的甘草酸浓度相似甚至更高,并证实在受控环境下,可以在短期内实现高浓度的甘草酸生产[17]。
UV-B可诱导产生更多的多酚类化合物,而黄酮类化合物是其中最大的类别。睡茄在UV-B辐射下叶片和根部4-香豆酸Co-A连接酶(4CL)、肉桂醇脱氢酶(CAD)、查尔酮异构酶(CHI)、苯丙氨酸解氨酶(PAL)和二氢黄酮醇还原酶(DFR)的活性增加[18]。与此相似,UV-B辐射显著提高了辣椒叶片中绿原酸和芹菜素8-C-己糖的含量,且苯丙氨酸解氨酶(PAL)和查尔酮合成酶(CHS)基因的表达量在UV-B处理下也有所增加[19]。
-
UV-C照射几乎可以诱导和提高所有酚类化合物中类黄酮的生物合成[9]。最近的一份研究表明,金荞麦在光周期为16L/8D h的UV-C(5.4 kJ/m2)处理后,积累了最优的生物量(438.3 g/L 鲜重,16.4 g/L 干重)、总黄酮含量(TFC: 4.05 mg/g)和酚类物质含量(TPC: 11.8 mg/g)[20]。虎杖在UV-C(10 W/m2)照射6 h和12 h后,白藜芦醇含量分别比未处理的显著提高了2.6倍和1.6倍。UV-C处理后,苯丙氨酸解氨酶、肉桂酸4-羟化酶、4-香豆酸辅酶a连接酶和二苯乙烯合酶(STS)等直接参与白藜芦醇合成的基因被强烈诱导,并确定了MYB、bHLH和ERF家族中的几个成员作为白藜芦醇生物合成基因的潜在调控因子[21]。
-
红光和蓝光较易被叶绿素吸收,所以是调节药用植物SMs含量的重要光质。Kapoor等[22]研究发现,蓝光处理的愈伤组织培养至第21天,积累的红景天苷(3.12 mg/g 干重)量最大,TFC(5.53 mg/g 干重)和总酚(TPC,11.84 mg/g 干重)的积累量均高于其他光照条件,但红光下培养的愈伤组织在第21天生物量(7.43 g/L)最大,红光培养下愈伤组织的比生长率最高(0.126/d),倍增时间最高(132.66 h),即红光刺激最大生物量积累,蓝光促进红景天苷、酚类物质和黄酮类物质积累。Manivannan等[23]研究发现,不同光照处理丹东玄参时,TPC和TFC的含量在蓝光处理下显著高于红光处理。值得注意的是,蓝光显著提高了TFC(比冷白色荧光灯组高65.68%)和TPC(比冷白色荧光灯组高55.14%)的含量。在对墨旱莲的研究中,红光处理可促进酚类物质(57.8 mg/g)和黄酮类物质(11.1 mg/g)的产生。蓝光处理下,4种主要化合物香豆素(1.26 mg/g)、小泡蛋白(5.00 mg/g)、威地洛内酯(32.54 mg/g)和去甲基威地洛内酯(23.67 mg/g)以及2种次要化合物β-香树素 (0.38 mg/g)和木犀草素(0.39 mg/g)的产量均有所增加[24]。Nadeem等[25]研究了发光二极管对罗勒愈伤组织中生物活性成分积累的影响,发现各处理中,蓝光处理下愈伤组织TPC含量最高,红光处理下愈伤组织TFC含量最高。高效液相色谱法(HPLC)分析表明,蓝光条件下迷迭香酸(96.0 mg/g 干重)和丁香酚(0.273 mg/g干重)浓度最高,分别为对照组的2.46和2.25倍。红光下愈伤组织中花青素(0.122 mg/g 干重)和芍药苷(0.127 mg/g 干重)含量最高。连续白光下培养的愈伤组织中,氨基酸积累量为81.40 mg/g(干重),是对照的4.52倍。水飞蓟是一种被广泛研究的著名保肝药材。研究发现红光能显著提高该植物的酚类物质、黄酮类物质和超氧化物歧化酶活性。在红光下,HPLC分析显示水飞蓟素总浓度(18.67 mg/g 干重)是对照的2倍。当暴露在红光下时,水飞蓟素、异水飞蓟素、水飞蓟宾A、水飞蓟宾B和水飞蓟宁的含量都很高[26]。此外,蓝光对远志根中TFC和TPC积累有促进作用[27],可有效促进半夏块茎中总生物碱的累积[28],也可以促进淫羊藿苷类黄酮的形成[29]。但也有相反的情况,如在对绞股蓝的研究中,红光比蓝光更有利于绞股蓝总皂苷的积累,但是红光却抑制绞股蓝TFC的积累[30];红光处理下黄花蒿的青蒿素和青蒿酸含量较低[31]。
也有研究表明,红蓝光组合能更好的促进药用植物中SMs的积累。李琪等[32]研究表明,与单色红光和蓝光相比,红蓝复合光更有利于金线兰组培苗的生长发育以及SMs的积累。其中,红蓝(1∶1)组相较于其他组合光更能显著促进金线兰黄酮类、酚类、氨基酸类等多种次级代谢产物的积累。因此,红蓝(1∶1)可以作为金线兰组织培养的最佳光质。刘建福等[33]发现,R7B3时最有利于姜黄根茎SMs的生成,此条件下,姜黄素类化合物含量达到最大值, 去甲氧基姜黄素、姜黄素和双去甲氧基姜黄素含量分别比白光(对照)显著提高44.32%、77.39%和43.80%。
-
有研究表明黄光可以促进SMs的产生。黄光处理下拟巫山淫羊藿的黄酮类化合物含量高于红光、蓝光和白光处理。RNAseq和qPCR分析表明,黄光可显著上调黄酮类生物合成基因CHS1、F3H1、PT_5和raGT_5的表达水平,可能是黄酮类生物活性物质积累增强的原因之一。耐阴植物通常生活在阳光较少的地方(如林下作物),光合作用驱动的波段,如红光和蓝光优先被上层冠层过滤掉,可利用的有效辐射主要为绿光、黄光和红外光。因此,拟巫山淫羊藿可能是为了适应遮荫条件下有限的黄色辐射光谱而进化的[34]。
-
研究表明绿色植物之所以呈现绿色是由于绿光大部分被植物叶片反射而很少被叶绿体吸收,因此绿光也被认为是生理无效光[7]。确实有很多研究表明绿光不利于SMs的积累。如绿光对绞股蓝总皂苷、TFC的积累有明显的抑制作用[30]。但也有研究表明绿光由于其高的透过率和反射率,可以深入叶片内部参与并影响光合作用,特别是在强烈的白光下,较低的叶绿体吸收任何额外的绿光都会比额外的红光或蓝光更大程度地促进叶片的光合作用[35]。青钱柳是一种兼具经济价值和药用价值的树种。研究发现与白光、蓝光和红光相比,绿光处理下青钱柳叶片TFC含量最高[36]。此外,绿光虽然抑制半夏块茎中总生物碱的积累,但却促进了黄酮类物质的产生[28]。
-
近红外光是可以通过对光敏色素作用来直接调控植物的生长发育。低R/ FR条件会使拟南芥的叶片大而厚、生物质数量大,冷适应能力强[37]。在川贝母中异甾体类生物碱的合成中,也发现了近红外光具有类似的作用[38]。光敏色素控制植物的开花主要取决于红外光吸收型(Pfr)/红光吸收型(Pr)的比值,因此可以通过红外光照射来控制Pfr/Pr值,从而较精确地控制开花的周期,这对于以花入药的药用植物具有重要意义[39]。
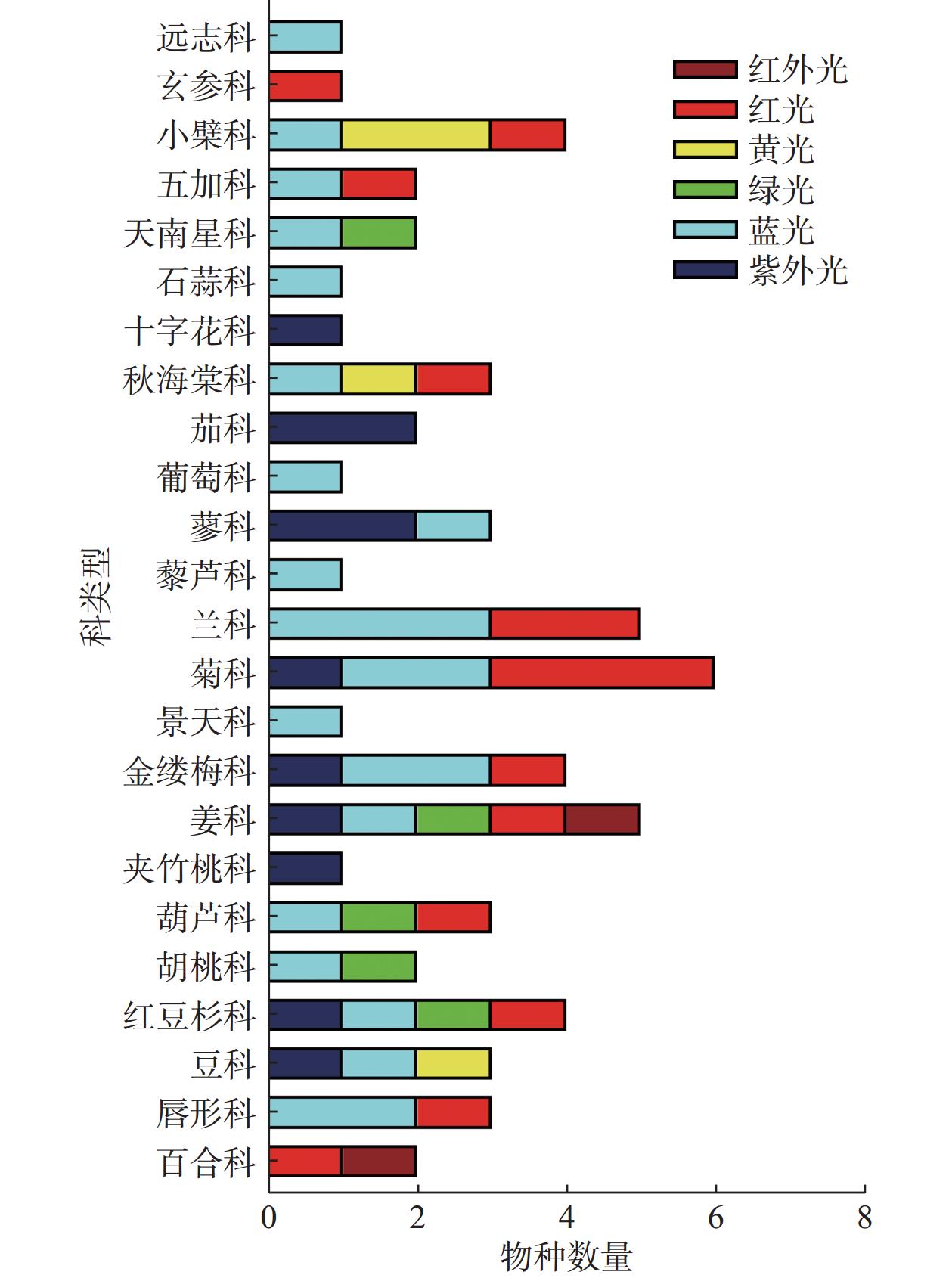
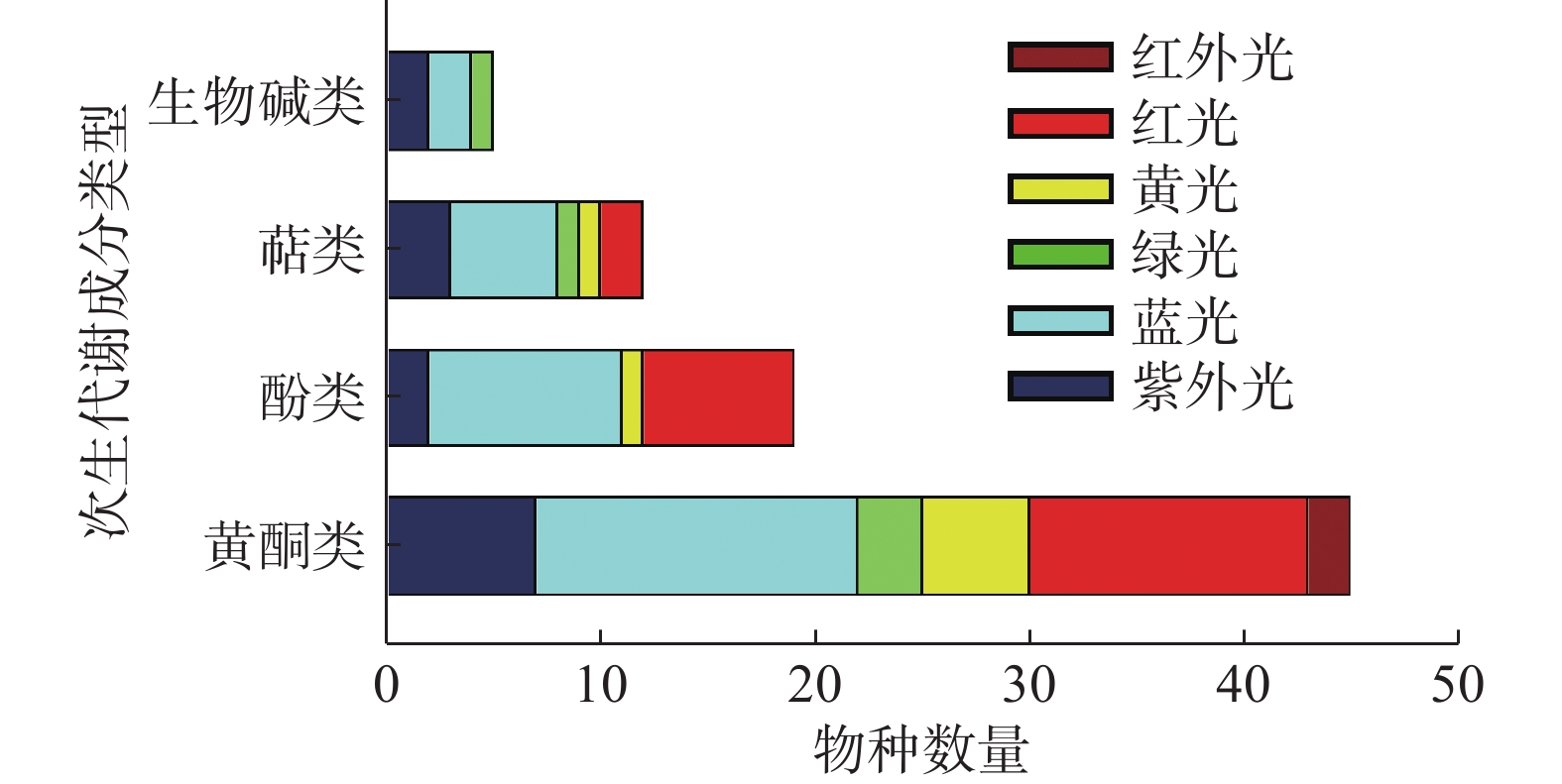
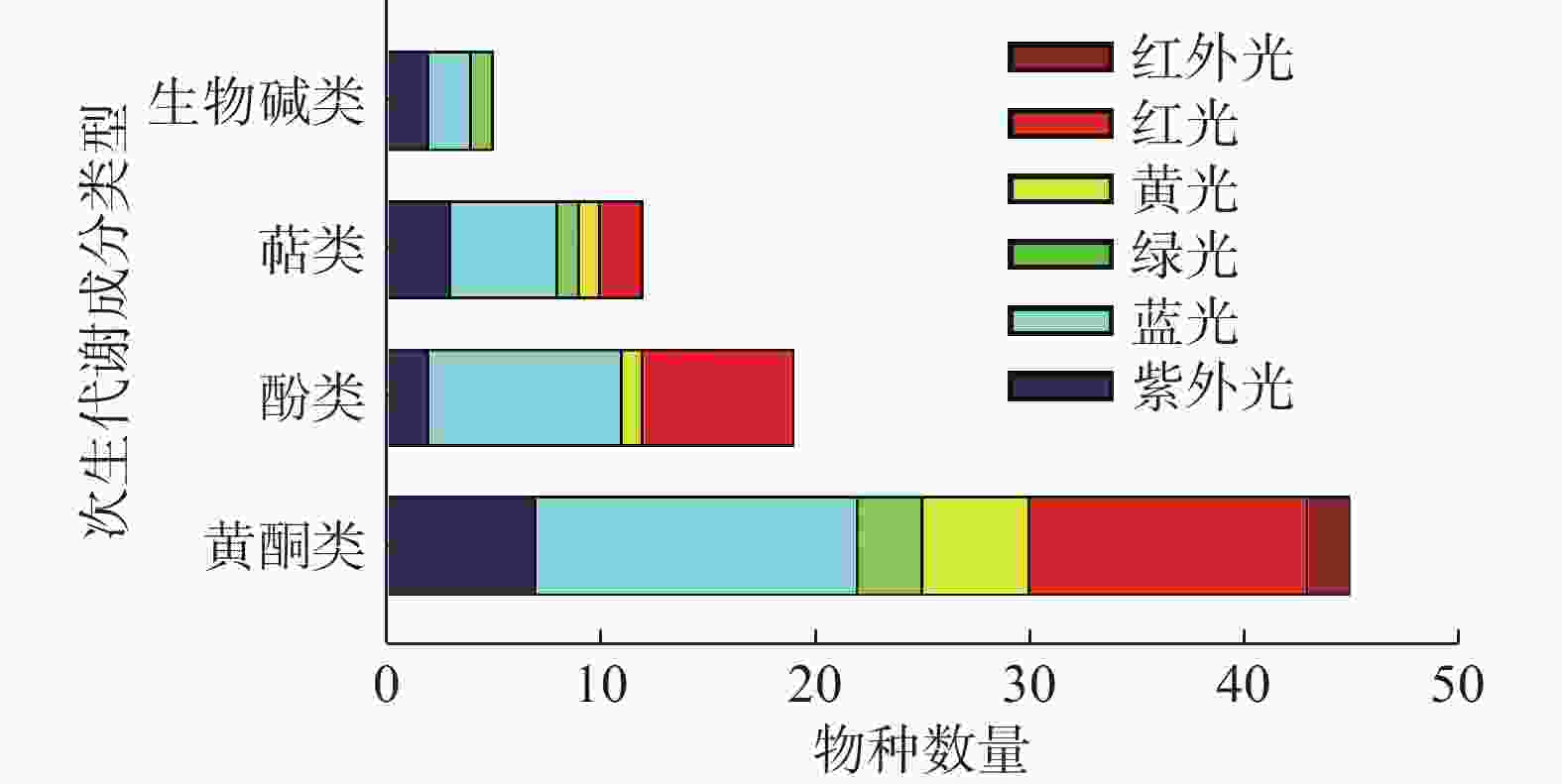
综上所述,不同的光质对不同植物甚至是同一植物的不同类型的SMs影响都是不一样的(见表1)。不同分类条件下光质对药用植物产生的影响见图2、图3所示,不同波长的光皆有可能成为诱导药用植物SMs积累的潜在因子。目前应用红蓝光调控黄酮类SMs研究较多,一方面是因为红蓝光较其它光质更易获取与利用,另一方面因为黄酮类次生代谢成分在药用植物中相对普遍存在,生物合成途径较为明确。不同光质对不同药用植物、不同的SMs存在不一样的调控作用,明确这些作用需要大量的实验去探索。UV在过去很长的一段时间里被视为不利于植物生长的有害因素,尤其是UV-C更被视为“灭生性辐射”,但现在许多研究表明低剂量的(适宜的)紫外辐射会触发植物体内抗逆境基因的表达,从而促进SMs的积累。
科名 拉丁名 中文名 光质 光强 作用时间 SMs影响 类型 参考文献 百合科 Fritillaria cirrhosa 川贝母 R
FR56 μmol/ (m2·s) 16 h
90 d异甾体类生物碱↑ 生物碱类 [38] 唇形科 Ocimum basilicum 罗勒 B 40~50 μmol/ (m2·s) 24 h
28 d迷迭香酸↑
丁香酸↑
总酚↑酚类 [25] 唇形科 Salvia miltiorrhiza 丹参 B
R300 μmol/ (m2·s) 16 h
20 d丹酚酸B↑ 酚类 [40] 豆科 Scutellaria baicalensis 黄芩 W 90 μmol/ (m2·s) 16 h
14 d黄芩苷↑
汉黄芩苷↑
黄酮类↑黄酮类 [41] 豆科 Astragalus membranaceus 黄芪 B
Y透光率85% 120 d 总皂苷↑
总黄酮↑萜类
黄酮类[42] 豆科 Glycyrrhiza uralensis 甘草 UVB 1.43 W/m2
0.43 W/m23 d
15 d甘草甜素↑ 萜类 [17] 红豆杉科 Taxus cuspidata 东北红豆杉 R
B
G40~50 μmol/ (m2·s) 15 d 总紫杉烷↑ 萜类 [43] 红豆杉科 Taxus cuspidata 东北红豆杉 UVB 3 W/m2 12 h 紫杉醇↑
类黄酮↑萜类
黄酮类[15] 胡桃科 Cyclocarya paliurus 青钱柳 B
G(800 ± 50) μmol/ (m2·s) 12 h
60 d山柰酚↑
异槲皮苷↑
槲皮素↑
总黄酮↑黄酮类 [36] 葫芦科 Gynostemma pentaphyllum 绞股蓝 B
R
G1500~2 000 Lx 12 h
60 d总黄酮↓ 黄酮类 [30] 夹竹桃科 Catharanthus roseus 长春花 UVB 1.26 W/m2 5 min 长春碱↑
长春新碱↑生物碱类 [14] 姜科 Curcuma longa 姜黄 G
FR280 μmol/ (m2·s) 13 h
60 d姜黄素类化合物↓ 黄酮类 [33] 金缕梅科 Loropetalum chinense 红花檵木 B+UVA
B60 μmol/ (m2·s) 12 h
15 d、30 d花色素苷↑
总黄酮↑黄酮类 [11] 景天科 Rhodiola rosea 红景天 B 25~27 μmol/ (m2·s) 16 h
30 d红景天苷↑
总黄酮↑
总多酚↑酚类
黄酮类[22] 菊科 Artemisia annua 黄花蒿 W
B(200 ± 10) μmol/ (m2·s) 16 h
7 d青蒿素↑
青蒿酸↑萜类 [31] 菊科 Eclipta prostrata 墨旱莲 R 40~50 μmol/ (m2·s) 24 h
28 d酚类↑
黄酮类↑酚类
黄酮类[24] 菊科 Silybum marianum 水飞蓟 R 40~50 μmol/ (m2·s) 24 h
28 d总水飞蓟素↑
总酚↑
总黄酮↑酚类
黄酮类[26] 菊科 Atractylodes lancea 茅苍术 9R1B 100 μmol/ (m2·s) 180 d 挥发油↑ 萜类 [44] 菊科 Artemisia annua 黄花蒿 UVB 1.5 μmol/ (m2·s) 4 h
9 d青蒿素↑
类黄酮↑萜类
黄酮类[16] 兰科 Anoectochilus roxburghii 金线莲 R1B1 (16± 1) μmol/ (m2·s) 12 h
120 d酚类↑
黄酮类↑酚类
黄酮类[32] 兰科 Arundina graminifolia 竹叶兰 1R3B
B45 μmol/ (m2·s) 12 h
40 d总酚↑
类黄酮↑酚类
黄酮类[45] 藜芦科 Paris polyphylla 陕产重楼 B 80 μmol/ (m2·s) 12 h
30 d总皂昔↑ 萜类 [46] 蓼科 Reynoutria japonica 虎杖 B 3000 Lx 30 d 白藜芦醇↑ 酚类 [47] 蓼科 Fagopyrum dibotrys 金荞麦 UVC 5.4 KJ/m2 30 min 总黄酮↑
多酚类↑酚类
黄酮类[20] 蓼科 Reynoutria japonica Houtt. 虎杖 UVC 6 W/m2 10 min 白藜芦醇↑ 酚类 [21] 葡萄科 Tetrastigma hemsleyanum 三叶青 B 1630~1660 Lx 12 h
30~45 d总黄酮↑ 黄酮类 [48] 茄科 Withania somnifera 睡茄 UVB 3.6 KJ/ (m2·d) 4 h 生物碱↑
黄酮类↑生物碱类
黄酮类[18] 茄科 Capsicum annuum 辣椒 UVB 80 W/m2 4 h 黄酮类↑ 黄酮类 [19] 秋海棠科 Gynura bicolo 紫背天葵 7R2B1Y (350±5) μmol/ (m2·s) 16 h
60 d总酚↑
类黄酮↑
花青素↑酚类
黄酮类[49] 十字花科 Brassica rapa 津田芜菁 UVA 3 W/m2 24 h 花青素↑ 黄酮类 [12] 石蒜科 Lycoris radiata 石蒜 B 50 μmol/ (m2·s) 12 h
30 d生物碱↑ 生物碱类 [50] 天南星科 Pinellia ternata 半夏 B 40 μmol/ (m2·s) 12 h
45 d总生物碱↑ 生物碱类 [28] 五加科 Eleutherococcus senticosus 刺五加 B
R透光率100% 20 d 总黄酮↓ 黄酮类 [51] 小檗科 Epimedium
pseudowushanense拟巫山淫羊藿 Y (18.2 ± 2.5) μmol/ (m2·s) 16 h
30 d黄酮类↑ 黄酮类 [34] 小檗科 Epimedium
pseudowushanense拟巫山淫羊藿 B
Y
R(18.2 ± 2.5) μmol/ (m2·s) 16 h
30 d淫羊藿苷类黄酮↑ 黄酮类 [29] 玄参科 Scrophularia kakudensis 丹东玄参 R 50 μmol/ (m2·s) 16 h
28 d总黄酮↑
总多酚↑酚类
黄酮类[23] 远志科 Polygala tenuifolia 远志 B 100 μmol/ (m2·s) 12 h
30 d总酚↑
总黄酮↑酚类
黄酮类[27] 注:B.蓝光;R.红光;Y.黄光;G.绿光;FR.红外光;UV.紫外线;↑表示SMs含量增加;↓表示SMs含量减少 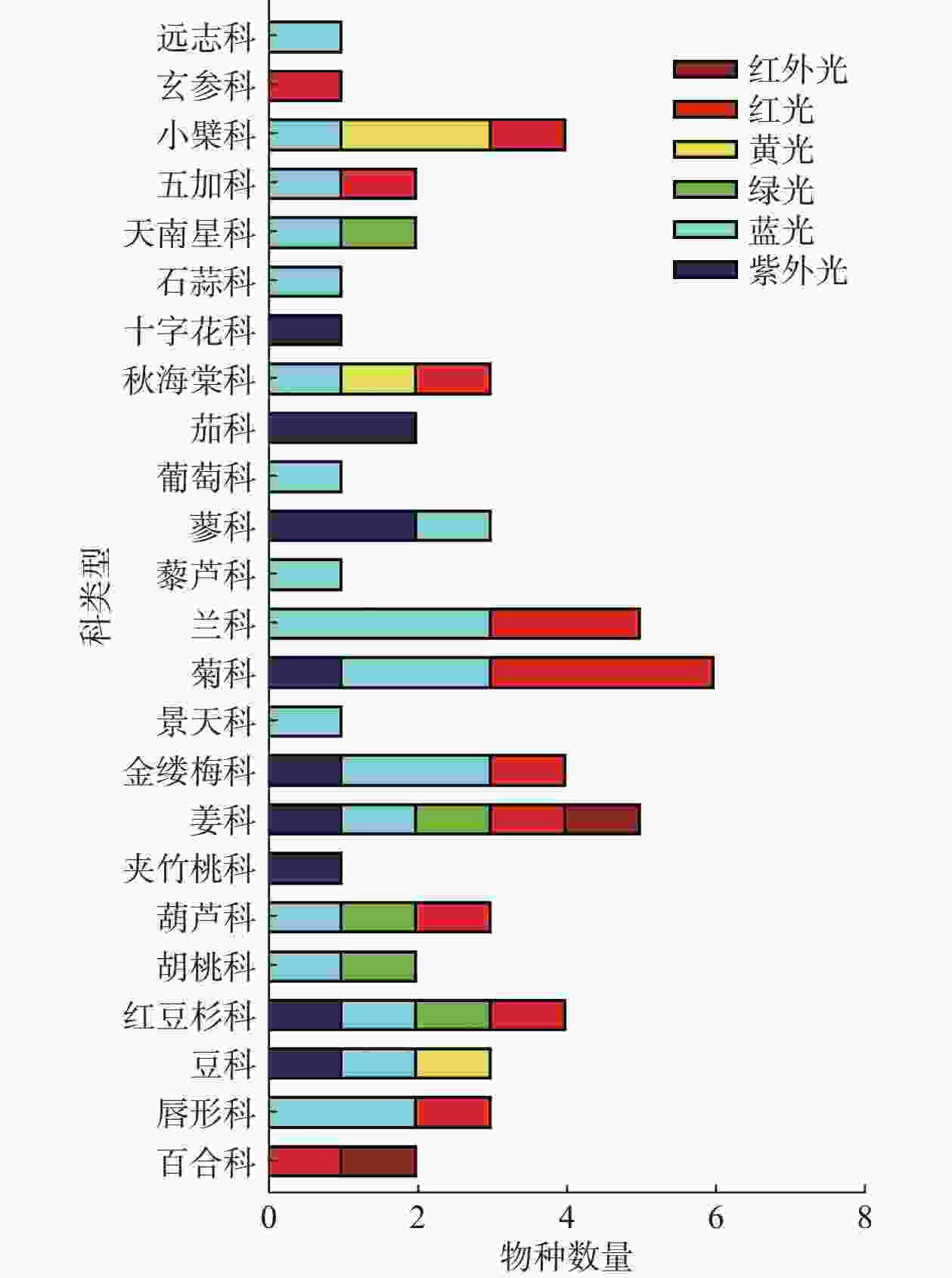
-
与其它生理过程相似,药用植物的次生代谢过程也受光照强度的影响。每种植物都有其适宜的光照强度,在其合适的光强下,药用植物才能更好的生长,产生更多的具有药理活性的SMs(见表2)。如典型的阳生植物金银花在阳光充足的地方品质更佳,Fang等[52]研究发现,完全光照促进了金银花中木犀草苷的生物合成和积累,与50%、25%光强相比,100%光照强度下参与黄酮类代谢的光响应转录因子(NAC、WD40、MYB、ERF、WRKY)在金银花中大量上调。野菊为喜光植物,有效成分含量如绿原酸、木犀草苷、蒙花苷、TFC等随光强升高而增加。钩藤虽也为喜阳植物,但在幼苗期也需要适度的遮荫处理。钩藤在70%~75%光强下适宜幼苗生长和生物量积累,在30%~35%光强下适宜钩藤幼苗有效成分(钩藤碱、异钩藤碱)的积累[53]。而对于阴性植物而言,需要在弱光条件下才能更好的生长。典型的阴生植物如滇重楼、三七、细辛等在栽培过程中都需要进行适度的遮荫处理或者种植在林下等潮湿背阴处。陈黎明等[54]研究表明,在透光率为16%的光强下,三七的光合作用效率最高,并且地下部分三七总皂苷含量达到最大值为4.75%。张勤涛等[55]发现,滇重楼在黄光处理下的最适光强是(75~100) μmol/(m2·s),而在蓝光处理下(100~150) μmol/(m2 ·s)[56],表明不同光质下植物的最适光强范围会有所差异。总之,不同植物对光照强度的需求不一,甚至在不同的生长阶段对光照强度的需求也会有所差别,需要充分考虑不同植物的生态习性从而做出有益的处理。
科 拉丁名 物种 生态习性 光质 不同光强 最适光强 SMs 参考文献 菊科 Aster indicus 马兰 喜阳 自然光 100%、62.29%、35.17% 叶中呈下降趋势
根中呈上升趋势
茎中62.29%最高黄酮类 [57] 菊科 Viola philippica 紫花地丁 中性 自然光 100%、80%、50%、35%、5% 35%~100%增加
但无显著性差异黄酮类
香豆素[58] 菊科 Chrysanthemum indicum 野菊 喜阳 自然光 100%、80%、60%、40%、20% 呈下降趋势60%较好 总黄酮
蒙花苷
木犀草苷
绿原酸[59] 蓝果树科 Camptotheca acuminata 喜树 喜阳 白光 800、500、300 μmol/ (m2·s) 300 μmol/ (m2·s) 喜树碱 [60] 藜芦科 Paris polyphylla 滇重楼 喜阴 黄光 100、75、50、25 μmol/(m2·s) 75~100 μmol/ (m2·s) 皂苷类 [55] 藜芦科 Paris polyphylla 滇重楼 喜阴 蓝光 200、150、100、50 μmol/ (m2·s) 100~150 μmol/ (m2·s) 皂苷类 [56] 马兜铃科 Asarum heterotropoides 北细辛 喜阴 自然光 100%、50%、28%、12% 50% 挥发油
细辛脂素[61] 木樨科 Syringa oblata 紫丁香 喜阳 自然光 100%、50%~60%、25%~35% 50%~60% 芦丁 [62] 茜草科 Uncaria rhynchophylla 钩藤 喜阳 自然光 100%、70%~75%、
30%~35%、5%~10%前期70%~75%
后期30%~35%生物碱 [53] 忍冬科 Lonicera japonica 金银花 喜阳 自然光 100%、50%、25% 100% 木犀草苷 [52] 伞形科 Changium smyrnioides 明党参 中性 自然光 100%、60.54%、44.84%、31.39%、10.56% 31.39%~60.54% 甘露醇
胆碱[63] 五加科 Panax notoginseng 三七 喜阴 自然光 32%、16%、8%、4%、2% 16% 总皂苷 [54] 远志科 Polygala tenuifolia 远志 喜阳 白光 500、300、100 μmol/ (m2·s) 无显著性差异 总黄酮
总酚[27] -
光周期即昼夜的相对长度,根据植物开花对日照长度的反应可分为:长日照植物、短日照植物和中日照植物。合适的光周期可以促进SMs的积累(见表3)。朱宁等[64]研究了不同光周期对龙眼胚性愈伤组织的影响,发现在短光照处理下(6 h/18 h L/D)类黄酮的含量和产量最高;全光照处理下(24 h)类胡萝卜素含量和产量最高。Tusevski等[65]发现贯叶连翘毛状根在光周期16 h/8 h下可以从头合成五种氧杂蒽酮、三种黄酮苷和两种酚酸,但在暗培养下更有利于黄烷-3-醇的合成。刘静等[66]的研究表明,在光周期12 h/d处理下,明显促进醉马草内生真菌共生体幼苗麦角酰胺和麦角新碱的积累。而朱娇等[67]的研究表明,不同光周期对西红花的品质没有影响,但在光周期8 h/16 h下,有利于西红花生长、促进初期提前开花。由此可见,光周期可以调控药用植物中SMs的产生和积累。
科 拉丁名 物种 光强 不同光照时长 最适光照时长 SMs 参考文献 禾本科 Achnatherum inebrians 醉马草 2500 Lx 16、14、12、8 h/d 12 h/d 麦角酰胺
麦角新碱[66] 金丝桃科 Hypericum perforatum 贯叶连翘 50 μmol/(m2·s) 16、0 h/d 16、0 h/d 酚酸、黄酮醇、氧杂蒽酮(16)
黄烷-3-醇(暗培养)[65] 落葵科 Basella alba 落葵 45 μmol/(m2·s) 24、16、0 h/d 16 h/d 总酚
黄酮类[68] 无患子科 Dimocarpus longan 龙眼 36 μmol/(m2·s) 24、18、12、6、0 h/d 24 h/d
6 h/d类胡萝卜素(24)
类黄酮(6)[64] 鸢尾科 Crocus sativus 西红花 (83±8) μmol/(m2·s) 14、12、10、8 h/d 8 h/d 西红花苷(无影响) [67] -
植物对光的响应具有特异性,光对不同植物SMs生物合成的影响不同。在植物光调控领域,光质、光强、光周期实际上是相互影响的,改变其中任何一个因素得到的实验结果可能会有所不同。在光强为(18.2 ± 2.5) μmol/(m2·s)时,黄光、蓝光和红光下淫羊藿苷类黄酮含量均高于白光;而在(90.9 ± 2.5) μmol/(m2·s) 光强下,白光促进淫羊藿苷类成分合成的效果却强于黄光、蓝光和红光[29]。再如,滇重楼在蓝光下得出的最适光强为(75~100) μmol/(m2·s);而在黄光下得出的最适光强却是(100~150) μmol/(m2·s)。光敏色素、隐色素、向光素、蓝光受体、紫外线感光器是目前已鉴定出的五类光感受器,光照通过与光感受器作用,调控SMs生物合成中涉及的转录因子或关键酶的基因表达。例如,当植物暴露于UV-B辐射时,UVR8同二聚体经历单体化并与COP1相互作用,从而增加COP1的稳定性并诱导HY5的表达,HY5结合一些转录因子基因的启动子,如AaWRKY9、GLANDULAR trichomspecific WRKY 1 (AaGSW1)和AaORA,从而上调它们的表达,这些转录因子刺激相关生物合成基因的表达,提高青蒿素的含量[69-70]。植物不同的生长时期对光的需求是不一样的,即植物对光的需求具有动态性,但大多数研究给予的光照条件却是静态性的。因此,目前许多对于光调控药用植物SMs的研究是有局限的,即如对光的某一方面(光质或光强或光周期)进行考察,得出的实验结果可能具有片面性,如能对光相互影响的三个方面(光质、光强、光周期)同时进行综合考察有可能得出较优的光配方。
体外培养(如组织培养)生产具有药理活性的SMs可以对光照条件进行更为精准的调控。对于次生代谢工程而言,通过光调控提高SMs的产量和质量是一种比较高效的、经济的手段。目前药用植物绝大多数是在室外种植,人们对光的调控是受限的。因此,在进行实验时,应更多的考虑室外大规模种植时可利用的光调控手段进行研究,这样得出的科研成果才有可能投入实际的生产中。
药用植物中SMs是其主要的药效物质基础和品质内涵,药材内在品质主要取决于这类产物的合成与积累。在自然界中,光往往与其它环境因子(如温度、水分、微生物等)协同或拮抗地调控药用植物SMs的生物合成。某种植物中SMs的种类及合成量的阈值由自身的遗传特性决定,光与其他环境因素则对这些产物的表达量起调控作用。随着人们对天然活性产物需求的增加,许多可人为控制的光调控系统已被用于药用植物的栽培。光对植物的调控作用是复杂的,目前人们对完整的光信号通路仍然不清楚,可以结合转录组学、基因组学、蛋白质组学、代谢组学等多种组学方法,更加全面的了解光对药用植物中SMs的影响机制,为提高SMs的产量和质量提供新的思路和方法。
Effects of light regulation on the synthesis of secondary metabolites in medicinal plants
doi: 10.12206/j.issn.2097-2024.202305027
- Received Date: 2023-05-17
- Rev Recd Date: 2023-09-14
- Available Online: 2024-02-20
- Publish Date: 2024-02-25
-
Key words:
- light quality /
- light intensity /
- photoperiod /
- medicinal plants /
- secondary metabolites
Abstract: Secondary metabolites of medicinal plants are extremely important to human health because of their special pharmacological activities or efficacy. They are the main source of drugs, health care products, and cosmetics. As human beings continue to pursue health and longevity, the demand in the pharmaceutical market continues to grow. It becomes especially important to improve the production and quality of secondary metabolites of medicinal plants. Plant secondary metabolites are a kind of adaptation of plants to their environment and are the result of the interaction between plants and biotic and abiotic factors during the long-term evolution process. The production and accumulation of secondary metabolites in medicinal plants are mainly affected by plant genetic factors and environmental factors. Among them, light environment is extremely important for their synthesis. Therefore, light regulation has long been a research focus for many scholars in China and abroad. In this article, we the recent research progress on the effects of light regulation on the secondary metabolites of medicinal plants were reviewed, mainly focusing on the effects of light quality, light intensity and photoperiod, in order to provide theoretical basis and practical guidance for the efficient production of secondary metabolites with important pharmacological activities.
| Citation: | ZHANG Wei, MENG Xiangqing, SU Xiaohui, WANG Jinyi, LI Lihua, JIA Min. Effects of light regulation on the synthesis of secondary metabolites in medicinal plants[J]. Journal of Pharmaceutical Practice and Service, 2024, 42(2): 50-59. doi: 10.12206/j.issn.2097-2024.202305027

|
 DownLoad:
DownLoad: