




-
神经炎症是外周血液系统和中枢神经系统中免疫细胞活化和浸润、神经胶质细胞活化和炎性介质产生的过程[1]。脊髓中产生的炎性神经胶质介质调节突触传递,诱导和维持慢性疼痛,并提供神经炎症和慢性疼痛之间的联系[2]。炎症疼痛是一种常见的慢性疾病,以往研究提示,前动力蛋白(prokineticin,PK)系统参与组织损伤和神经损伤后的外周和中枢敏化,其在炎症疼痛中的作用已经被证实[3]。本文针对近年发表的有关PK系统参与炎症疼痛的研究文献,回顾性分析该镇痛靶点所发挥的镇痛机制。
HTML
-
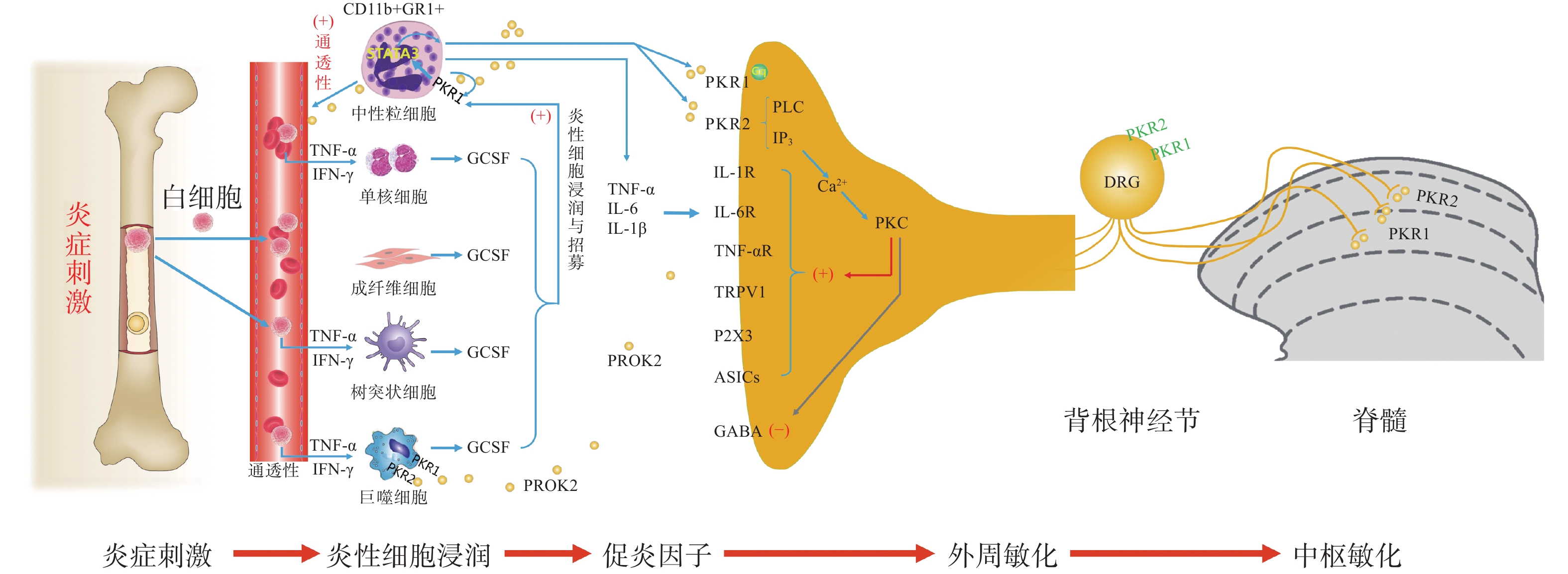
PK信号通路是近年来新发现的介导疼痛发生和维持的重要调节通路,包括两种结构上关联的小分子肽PK1和PK2,以及相应的G蛋白偶联受体PKR1和PKR2[4],PKs及其受体广泛分布在许多人体组织中,例如卵巢、睾丸、肾上腺、胎盘、子宫、大脑、肠道、心脏、骨髓和外周血。虽然PKR1和PKR2在多种组织中共同表达,但PKR1主要表达于外周组织,包括生殖系统[5]的内分泌腺和器官、胃肠道、脾脏、胰腺、肺、心脏和血细胞[6]。PKR2主要在脊髓背角神经元和星形胶质细胞中表达,PKR1主要存在于星形胶质细胞和外周神经元的末端,主要分布于背角的表层(图1)。PK2对这两种受体的亲和力均略高于PK1,而PK1在DRG(背根神经节)和脊髓中不表达,更多地参与血管生成[7],PK1是强有力的血管生成因子,被认为在内分泌腺、心血管、肾脏、肿瘤等血管新生中发挥重要作用。
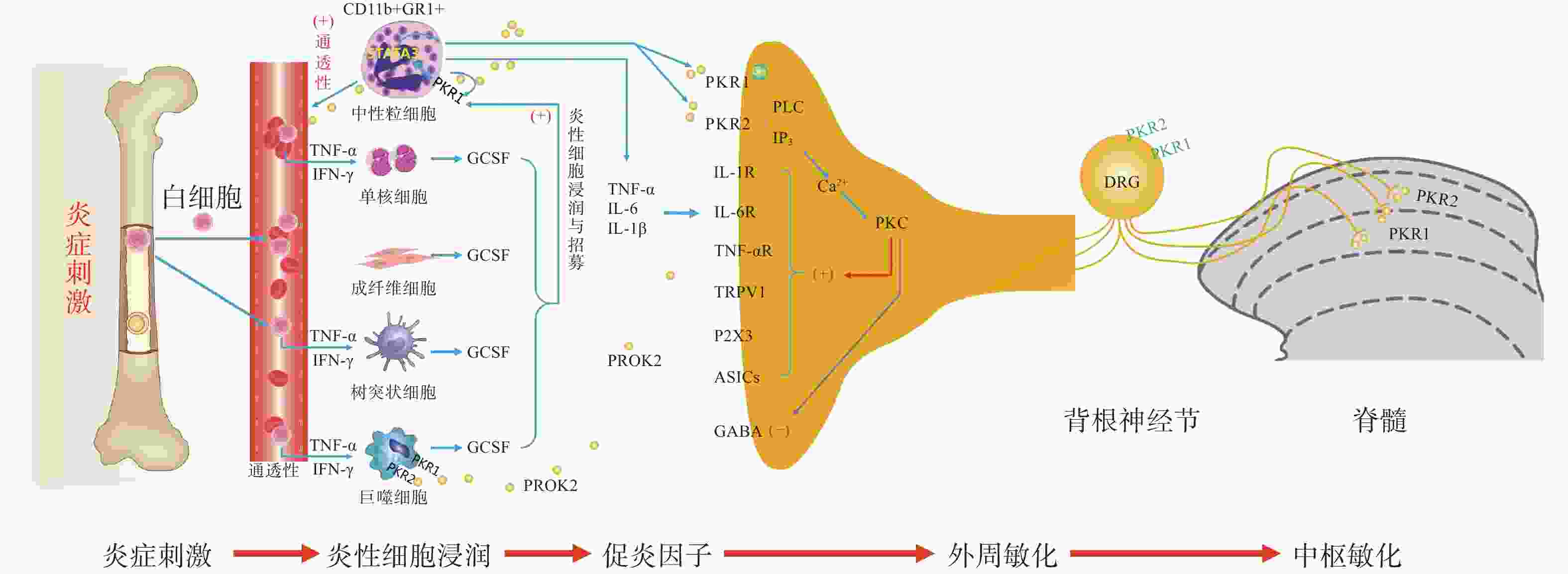
PKs在神经系统、免疫系统、生殖系统、心血管系统等多部位发挥广泛的生理作用[8],此外PK2可通过降低对化学和机械刺激的疼痛阈值,PKRs的拮抗剂可以用来缓解关节炎炎症和急慢性疼痛[9-10]。PKs通过激活单核细胞、巨噬细胞和树突状细胞上的PKRs,释放炎性细胞因子,触发和维持炎性疼痛(图1),调节免疫炎症反应,而在外周,PK2 /PKR2参与心脏和肾脏新生血管的形成,PKR1的缺失会引起心脏和肾脏的结构和功能变化[11-12]。因此,PKs系统在体内发挥着广泛的生理作用,也是众多疾病的潜在治疗靶点。
-
PK系统在调节与伤害相关的伤害性事件方面起着关键作用,因为它可以调节炎症反应,并同时作用于中枢和外周神经系统。PKRs的激活可以引起痛觉感受,参与痛觉感受器对不同刺激的敏感性。PK系统(PKs和PKRs)是在免疫细胞中参与炎症发生和疼痛传递的重要环节,如PK2参与免疫调节,诱导骨髓细胞分化为单核细胞系和巨噬细胞系,并通过激活单核细胞、巨噬细胞和树突状细胞上的PKRs,释放IL-1β、IL-6、TNF-α等炎性细胞因子,触发和维持炎性疼痛,调节免疫炎症反应[13]。
-
PK2在炎症细胞中呈强上调,并在病理状态下维持炎症循环。在纳摩尔范围内,PKs结合并激活受体PKR1和PKR2,使PK2在粒细胞、树突状细胞和巨噬细胞中表达,尤其在局部炎症后表达增加。此外,PK2在免疫细胞和神经胶质细胞中被强烈上调,并在炎症组织中,维持促炎循环。在中性粒细胞依赖性炎症和超痛觉过敏中起关键作用[14]。第一个关于PK的促痛作用的证据来自于对啮齿动物全身注射Bv8/PK2,激活其位于痛觉传入通路的PKRs受体而引起机械和热刺激痛觉过敏[15-16]。在完全弗氏佐剂(CFA)诱导的慢性炎症动物模型中,炎症与浸润炎症组织的粒细胞中PK2的过表达呈高度正相关,而PK2的上调则与炎症相关的痛觉过敏也有关,PKR1拮抗剂PC1能够有效消除CFA引起的痛觉过敏,PC1能明显抑制疼痛、水肿和外渗[17]。PK2在人和动物炎症组织中过表达,主要在浸润的中性粒细胞中作为主要的前感受介质之一,通过激活初级传入神经元上的PKRs受体(图1),增强伤害性信号向中枢神经系统传递。PK2反过来调节血管生成和血管通透性,激活巨噬细胞,并通过PKRs调节毛细血管内皮细胞和白细胞的免疫反应,使得炎症反应循环。
PKR1和PKR2基因的缺失大大降低了炎症诱导的热敏感性和机械敏感性[18-19],但是只有PKR1基因的缺失才会降低PK2的上调,这表明这两种受体虽然都介导疼痛的感受和传递,但只有PKR1参与了炎症过程中Bv8/PK2表达水平的增强。外周局部给予PKRs拮抗剂(PC1或PC7)后疼痛明显减轻,与Bv8/PK2对血管通透性的影响效果一致。在CFA诱导的大鼠或小鼠炎症模型中,在它们健康的爪子中很难检测到的PK2 mRNA,而在炎症皮肤中显著增加,且与浸润细胞(粒细胞和巨噬细胞)相关,并与疼痛和其他炎症在时间上相关。粒细胞释放PK2直接作用于痛觉受器,调节急性炎症性疼痛,进而发挥趋化作用,诱导促炎性巨噬细胞表型,并使Th1(T淋巴细胞1)/Th2(T淋巴细胞2)失衡向Th1倾斜[20-21],抑制Th1介导的免疫反应发挥抗炎作用。
PK2通过激活初级传入神经元上的PKR1和PKR2参与调节痛觉感知(图1),PKRs根据细胞定位可能与G蛋白结合,并激活不同的细胞内信号通路。在单核细胞/巨噬细胞/小胶质细胞谱系中,以及在DRG及脊髓中,PKRs是Gq偶联受体,通过激活磷脂酶C-β (PLC)和形成IP3,促进细胞内Ca2+的动员,诱发蛋白激酶C (PKC) -ε易位到质膜,外周高度表达的PKRs能够对TRPV1(瞬时感受器电位香草素亚型1)和TRPA1 (瞬时感受器电位锚蛋白亚型1)的促进性激活有助于外周敏化[22],尤其是PKR1定位于表达TRPV1和TRPA1的伤害性感觉纤维,在外周DRG水平这些共定位表达,为通过激活PKC-ε的痛觉敏化的协同作用提供了解剖学基础。在大鼠初级感觉神经元中,PK2还通过PKC信号通路增强了门控离子通道电流、抑制GABA激活电流、敏化嘌呤核苷酸P2受体(P2X)[23]。对PK2反应的DRG神经元中50%也表达参与疼痛处理的神经介质,如降钙素基因相关肽(CGRP)和P物质,且在这些神经元暴露于PKs时释放这些神经肽,提示PKs不仅仅参与调节中枢疼痛机制[24]。
-
脊髓PK2主要存在于小鼠脊髓星形胶质细胞中[25],最大密度的PKRs位于背角[26],这表明这些受体可能参与了伤害性信号的中枢传递。非神经元细胞如星形胶质细胞、小胶质细胞等都有PKR1表达,此外,PKR1受体在人和啮齿类单核/巨噬细胞上也比PKR2更为丰富。免疫组化定位显示,PKR1与进入背角和外皮层的伤害感受器末端有关,PKR2存在于沿着浅背角的神经元胞体中,也有可能是投射神经元。PK2诱导脊髓背角和活化的星形胶质细胞中PKRs的激活,有助于中枢敏化和维持慢性痛、神经性疼痛,而且PK2作用于单核细胞、巨噬细胞和树突状细胞上的PKRs,还能诱导炎症因子和致痛因子的释放[27]。在慢些压迫性损伤(CCI)模型中发现,PK2在脊髓星形胶质细胞和初级感觉神经元中表达上调[26]。同时,PK2可抑制下丘脑、导水管周围灰质(PAG)脑区中脑啡肽 (ENK)阳性神经元的动作电位放电,表明PK2能够抑制这类细胞释放阿片样物质。通过配体和受体在神经元和星形胶质细胞中的不同分布,PK2既可以诱导星形胶质细胞激活,发挥星形胶质细胞分泌生长因子的功能,也可以促进脊髓神经元的兴奋性突触传递,增强中枢敏化。PK2小分子拮抗剂改善持续疼痛超敏反应的能力以及在“神经元-神经胶质”和“神经元-免疫细胞”相互作用中分子变化,表明了PK2信号在这种行为中的重要性(图1)。阻断PK受体的药物可能为治疗慢性疼痛状态提供一种新形式[28]。在啮齿动物中注射PK2可激活位于疼痛传递主要部位的PKRs受体诱导对机械和热刺激的痛觉过敏[15]。痛觉过敏的初始阶段是由对伤害感受器的局部作用引起的,痛觉过敏的第二阶段是由于中枢作用引起的[24],表明PKs /PKRs在中枢与周围部位的作用存在差异。在敲除PK2基因的小鼠中进行研究表明,PKs在疼痛感知中具有直接作用,与正常野生型小鼠相比,基因敲除的小鼠对温度和化学刺激的伤害感受降低[18-19]。
损伤部位异常增加的IL-6、IL-1β反过来促进PK2的释放,进一步促进外周免疫细胞的过度激活,加速外周敏化的形成和维持。在DRG和脊髓中,PKR通过Gq激活后增加细胞内钙和诱发蛋白激酶C (PKC) -ε易位到质膜,PK2能够敏化表达TRPV1和TRPA1的痛觉感受器,从外周组织、DRG和脊髓等部位发挥多途径的抑制PK系统的激活而发挥镇痛效应。
综上所述,PKs在疼痛的发生和维持中发挥了重要作用,其在调节与伤害相关的伤害性事件方面起着关键作用,在未来的研究中,有望以PKs信号通路为靶点研究出新的治疗药物。
 DownLoad:
DownLoad: